
1.氷期の大気中二
1. 氷期の大気中二酸化炭素濃度減少の謎
地球環境の詳細な変動記録は氷河の氷の中にひっそりと残されていました。ボーリングで氷床に穴を掘ってゆくと、深くなるにつれてより昔にできた氷が手に入ります。その酸素や水素の同位体比の分析から、氷ができたときの地表面の平均気温が推定できることは、1960年代からコペンハーゲン大学のダンスガードらの研究で知られていました。氷の中には気泡やダストも含まれており、それらの分析から昔の環境を読み取ることができるかも知れなません。まず、空気が閉じ込められているはずであるから、変質をうけないようにうまく取り出せれば大気組成の変動がわかるでしょう。しかし、わずかな気泡中の二酸化炭素やメタンなど気体成分については、まだその測定手法に問題がありました。1980年代に入ってようやくその記録を読み出すことに、フランスやスイスの研究者が成功し、まず産業革命前は大気中の二酸化炭素濃度は280 ppmであったのに対し、最終氷期には200 ppm前後で約80 ppmも低かったことが、南極バード基地の氷床コアの分析から明らかにされました。引き続いて、南極ボストーク基地の氷床コアからは、過去42万年にわたる大気中の二酸化炭素やメタンが気温と連動し、いつも氷期にそれらの濃度が低くなる周期的変動をしていることも示されました[1]。氷期には陸上植物が減少していたことは明らかなので、その分の炭素と大気から減少した二酸化炭素はいずれも海洋に吸収されていたはずです。このことは、深海底堆積物中の有孔虫の炭素同位体比が、氷期にδ13C値にして平均0.35‰ほど負の側にずれている(つまり陸上植物の軽い炭素が海洋に加わった)ことによっても支持されています。
では、どのようにして氷期の海が過剰の二酸化炭素を取り込んだのでしょうか。
2. 溶解ポンプ
その原因としてまず考えられることは、表層水温の低下による二酸化炭素の溶解度の増加です。目には見えませんが、大気と海洋表層の間では海面を通して常に気体の交換が行われています。
海水の場合、気体の溶解度は物理化学的平衡によって水温が下がるとわずかながら増加し、塩分が高くなると逆に減少します。氷期には海洋表層の水温が低かったので、大気中の二酸化炭素がより多く海に溶け込み、濃度を減少させたのかも知れません。このメカニズムを「溶解ポンプ」と言います。
その効果は、氷期の海洋環境がどんな状態であったかによって決まります。最終氷期の表面海水の温度は現在より2℃前後低かったことが、深海堆積物中の有孔虫化石などの豊富な研究から知られています。一方、大陸氷床が成長したことによって、海水準が約120 mほど低下しました。そのために減少した海水量から、増加した塩分も計算できます。その結果は、両方の効果が相殺して、大気中の二酸化炭素濃度を高々10 ppm程度減少させるにすぎないことが判明しました。溶解ポンプではとても約80 ppmもの減少を説明するのは無理です。したがって、海洋中の物質循環における生物的あるいは化学的変化など、他の要因を考えなければいけません。
図1は、北太平洋における溶存無機炭素濃度の鉛直分布をそれぞれの支配要因に分けて示したものです[2]。深層水中の濃度は表層水に比べて約20%過剰になっています。これは海洋表層で光を利用して植物プランクトンがつくった有機物や炭酸カルシウムが、死後表層から沈降し深層水中でバクテリアによって分解、または化学的に溶解することによります。つまり、微小な海洋生物は、表層水の溶存無機炭素を体内に取り込み深層へ運んで放出・貯蔵する、いわばポンプの役割を果たしていることになります。氷期には、これら生物の働きによって(たとえば、生物生産量や炭酸カルシウムの生成・溶解過程が変わるなどして)、現在より多量の炭素が主に海洋深層に保持されていたと考えられます。そのさまざまな可能性をつぎに考えてみます。
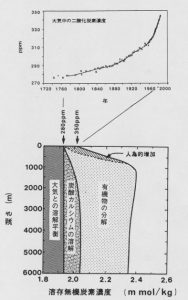
図1.北太平洋における溶存無機炭素の分布とその濃度を支配する因子
3. 生物ポンプ
海洋の生物生産は光の届く海洋表層(有光層)で行われるが、プランクトンは死後、有機物粒子として深海へ運ばれて表層から除かれるため、その不足した溶存無機炭素の補う形で大気から二酸化炭素が溶け込む。この過程を「生物ポンプ」と呼んでいる。その反応は

で示されます。海洋の生物生産を規制しているものは、主に微量必須栄養素である窒素やリンの濃度す。氷期の海水準低下で露出した陸棚の堆積物からリンが供給され、富栄養化して海洋の生物生産が増えたのではないかとする説もあります。つまり、過剰の栄養塩が海にもたらされた富栄養化現象です。また、海洋循環のモードが変わり、中・深層の海水に含まれた豊富な栄養塩が表層にもたらされて、光合成が活発になったとする説もあります。
海洋深層の流れは、グリーンランド東方海域や南極周辺海域の比較的塩分の高い表層水が冷却されて重くなるために沈み込むことで形成され、世界中の海を巡りめぐって、最終的には上昇して海洋表層にもたらされます。この一巡には、平均1000~2000年の時間がかかると言われています。中・深層の海水には、沈み込むときに海水が含んでいた分(約3分の1)と、後からプランクトンの死骸の有機物が分解して再生した分を合せて栄養塩が極めて豊富です。それらが表層にもたらされて植物プランクトンに有効に利用されれば、海洋の生物生産性は向上します。氷期には、図2に示すように海洋循環が変わっていたと想像されていますが、それに伴って中・深層の栄養塩がより有効に生物生産に組みいれられ、その結果、大気中の二酸化炭素濃度を減少させたとするモデルも相次いで発表されました。つまり、海洋中の栄養塩の総量は変わらなくても回転を速くし、生物ポンプの効率を上げることによって、大気中の二酸化炭素濃度を200 ppm以下にまで下げることも可能なのです。
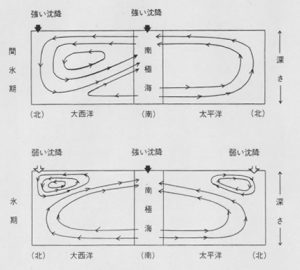
図2.氷期と間氷期の海洋循環の違い [2]
4. もう一つの微量必須元素:鉄
現在、中緯度海域の表層水は植物プランクトンに利用されて栄養塩がほぼ完全に枯渇していますが、リンや窒素が残っていながら生物生産があまり高くならない海域があります。東大平洋の赤道付近や南極周辺などが代表ですが、それらの海域では、最近微量必須元素の一つである鉄が不足しているために、生物生産が制限されていることが明らかになってきました。リービッヒの最少律で知られる必須栄養素の中に、カリウムやマグネシウムなどと並んで鉄があります。鉄はクロロフィルの構成元素で植物プランクトンの増殖に不可欠です。しかし、海水中では鉄は主に3価で存在し水酸化物を形成するため、非常に不安定で平均滞留時間が短いとされています。そのため、海水中に豊富なカリウムやマグネシウムとは異なり、鉄は測定するのが困難なほど濃度が低いです。カリフォルニアのモス・ランディング研究施設のジョン・マーチンがクリーン技術を駆使して分析した結果、表層海水中の鉄の濃度は1ナノモル(10^(-9)モル)以下であり、とくにリン酸や硝酸のようにとくに表層で低いことがわかりました。それらの海域で海水を採取し、少量の鉄を加えると活発に植物プランクトンが増殖することから、生物生産の制限要因となっていることが推定されました。
もし氷期にそのような海域に鉄が供給されたとすれば、生物ポンプの効率を高め、大気中の二酸化炭素濃度を減少させたことが期待されます。図3に示す氷床コア中のダストの量が氷期に激増したことでもわかるように、氷期には気温が低く乾燥気候だったため、土壌粒子などが舞い上がりやすい状態でした。そのダス卜に含まれた鉄が大気を通じて現在の富栄養低クロロフィル海域に運ばれ、海水に溶出して生物生産を高めたことが容易に想像されます。
しかしながら、これら生物ポンプのメカニズムの詳細はともかく、生物生産の変化に伴う有機物の深層へのフラックスの増加で、大気中の80 ppmの減少を説明しようとする試みには、どうしても越えられない大きなハードルがありました。それは、深層水中での有機物の酸化分解には溶存酸素が使われますが、あまりに有機物粒子の沈降が多くなると深層水の酸素が消費し尽くされて酸欠状態になってしまうということです。これは、深海の相当部分がヘドロ状態になることを意味し、氷期にも原生動物である底棲有孔虫が生きていたことを示す、深海堆積物の記録と明らかに矛盾します。
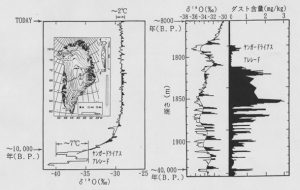
図3.グリーンランドのキャンプセンチュリー(左)とダイ3(右)の氷床コアの酸素同位体比およびダスト含量の変動記録
5. アルカリポンプの働き
そこで残る可能性は、炭酸カルシウムの生成と溶解のバランスが変わることによって、大気中の二酸化炭素が海に吸収されたのではないかとする考えです。二酸化炭素吸収の原理は中和反応で示され、溶存酸素は関係せず、アルカリ度が増加をします。したがってアルカリポンプと呼ばれますが、この過程は、深海が過剰の炭素を貯蔵しても無酸素状態にならずに済む今のところ唯一の解決策です。
海洋表層の海水は炭酸カルシウムに対して過飽和の状態にあり、有孔虫、円石藻、サンゴなどの生物が炭酸カルシウムを生成します。つまり、上記の反応が右から左へ進みます。一方、深海では圧力がかかり炭酸カルシウムの溶解度が増すことや有機物の分解のために二酸化炭素の分圧が高くなることから、ある深度を越えると未飽和になり、沈降してきたプランクトンの炭酸カルシウム殼は溶解します。表層海水のアルカリ度が氷期に高かったことは、二酸化炭素の大気と海水間の物理的な溶解平衡から計算で求めることが可能です。図4に示すように、最終氷期の表層海水は、産業革命前に比べてpHは0.15程度、またアルカリ度は110マイクロ当量ほど高かったことがわかります。そこで氷期には何らかの理由で、炭酸カルシウムがよく解けるようになったのではないかとする説が出されました。たとえばマサチューセッツ工科大学のE. A.ボイルによれば、生物生産が高くなって海底に到達する有機粒子のフラックスが増大し、その分解によって 生じた二酸化炭素が海底の炭酸カルシウムの溶解を加速することが考えられます。その結果、深層水のアルカリ度が増加し、その海水が海洋循環によって表層に出て大気に接すると、二酸化炭素を吸収することになります。具体的にその効果を論じた論文もその後いくつか発表されています。しかし、たとえこのように深海底で炭酸カルシウムの溶解が増えたとしても、その影響が大気に現れるには、海洋循環の時間スケールから考えて少なくとも数百年はかかるに違いありません。しかし、氷床コアの二酸化炭素濃度や泥炭コアの炭素同位体が示す大気中の二酸化炭素濃度の変動は、わずか20~30年で起っています。つまり、この深海底炭酸塩溶解説だけで説明するのには無理があるといえます。
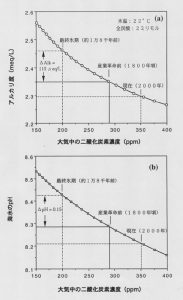
図4. 大気と平衡にある表層海水のアルカリ度(a)とpH(b)
6. シリコン仮説
このような状況の中で野崎は新しい解決策を思いつき、大場と共同で論文に発表しました[3]。つまり、深海での炭酸カルシウムの溶解が増えたのではなく、その殼をもつプランクトンの増殖が抑制されたのではないかと考えたのです。表層海水にリンや窒素など他の栄養塩があれば、溶存ケイ素がある場合にはオパール(無定形ケイ酸塩)の殼をもつ珪藻が優先的に繁殖することは、以前から知られていました。ケイ素が欠乏してなお他の栄養塩が残っている場合に、円石藻が増殖します。栄養塩の奪い合いでは、炭酸カルシウムの殻を持つ円石藻はなぜかケイ藻に引けを取るという自然界の摂理があります。そのような条件下では、円石藻は必要な窒素やリンを珪藻に奪われて増殖しにくいのではないかと考えたのです。氷期には海洋表層へ運ばれるエアロゾルの量が増大し、そこから溶け出した溶存ケイ素は珪藻の増殖を促進したはずです。
したがって、珪藻が卓越する期間が長くなるとか、あるいは珪藻が栄養塩を使い切ってしまって円石藻が成長できないなどの現象が起これば、(2) 式に照らしてアルカリ度は上昇し、大気から二酸化炭素を吸収できることになります。新たな補足を加えてその仮説を述べると次のようになります(図5参照)。
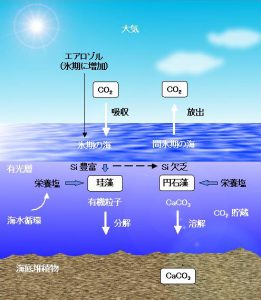
図5.シリコン誘導炭素循環モデルの概略
(1) カナダ北方の氷河崩壊(ハインリッヒ・イベントに対応)で北大西洋表層に淡水が流入し、熱塩循環(ブロッカーのコンベヤーベルト)を停止しそうなほど弱めました。その結果、北大西洋での表層水が停滞して沈降しなくなり、メキシコ湾流は北上できなくなってヨーロッパが急速に寒冷化します。
(2) 寒冷化によって、乾燥気候となった大地から土壌粒子が舞い上がりやすくなり、海洋表層へのエアロゾルの降下量が増加しました。つまり、エアロゾルによってもたらされたケイ素が海洋表層に豊富になり、珪藻の増殖を促進すると同時に、炭酸カルシウムの殻をもつ円石藻の増殖を抑制します。
(3) その結果、表層海水における炭酸カルシウムの生成量が相対的に減少し、沈降粒子の有機炭素/ 炭酸カルシウム比を押し上げます。そして海水のアルカリ度を増加させるとともに、pH が高くなって大気から二酸化炭素を吸収します。
このように海洋表層での過程ならば、20~30年で大気中の二酸化炭素濃度を変化させることも可能でしょう。生物の種の交代にはこのように一定のルールがある場合があります。この生物リンケージを通して、化学的には独立した反応である炭酸カルシウムの生成・溶解サイクルとシリカのサイクルが、自然界では巧妙に結びついているようです。
7. 新たな証拠探し
最近のモデル計算では、全海洋で生産される炭酸カルシウムが4割減少すれば、シリコン仮説のメカニズムで氷期大気の二酸化炭素濃度の説明が可能といわれています。円石藻と珪藻の種の交代は、リン、窒素、鉄などに対して溶存ケイ素の供給が相対的に不足した海域で実際に起こり得ます。北大西洋、赤道大平洋や南極海の南緯45~50度以北では、溶存ケイ素と硝酸の比が珪藻が必要とする1以下でその候補海域ということになります。最近、コロンビア大学ラモント地球観測研究所のC.D.チャールズらが南極周辺海域の深海堆積物の酸素同位体比とともにオパールと炭酸カルシウム含量を詳しく発表していますが、その一例を図6に示しました。堆積物中のオパール含量は、海水を沈降中あるいは海底で埋没するまでの間に溶解されずに、残ったほんの一部分にすぎないので、その溶解と保存に関する様々な過程が変われば影響されます。しかし、チャールズら[4] は、様々な検討を行った後、オパール含量は主に海洋表層での生物生産を表しているものと結論している。同様の仮定は、炭酸カルシウムについても成り立つでしょう。
図6から明らかなように、過去約1万年の間は炭酸カルシウムが卓越していますが、1万9千年から2万5千年の最終氷期の時代には、炭酸カルシウムは数%にまで後退し、珪藻が主になることがわかる。珪藻と円石藻の種の交代が起っていることは、図7に示すオパールと炭酸塩のきれいな逆相関関係からも推定できます。また、過去1万年の間は約90%が生物性炭酸塩とオパールで占められていますが、最終氷期には20~25%で、その他は陸から運ばれた粘土鉱物などです。堆積物の年代から陸起源微小粒子の堆積速度を計算すると、氷期の方が現在の間氷期より1桁大きいことが分かります。氷期に露出した陸棚から運ばれたものも含まれるかも知れませんが、大部分は大気を経由して運ばれたものと考えられます。
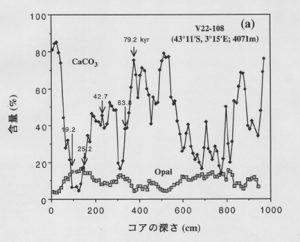
図6. 南大洋深海コアの炭酸カルシウムとオパール含量の変動[5]。図中の数値は千年の単位の年代を表す
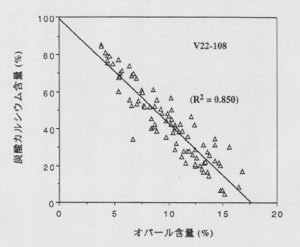
図7. V22-108コアの炭酸カルシウムとオパール含量の関係
参考文献:
[1] Petit J. R. et al. (1999), Climate and atmospheric history of the past 420,000 years from the Vostok ice core, Antarctica. Nature 399, 429-436.
[2] 野崎義行 (1994)、地球温暖化と海、東京大学出版会、p196.
[3] Nozaki, Y. and T. Oba (1995) Dissolution of calcareous tests in the ocean and atmospheric carbon dioxide, In: Biogeochemical Processes and Ocean Flux in the Western Pacific. H. Sakai and Y. Nozaki, eds., TerraPub., Tokyo, p83-92.
[4] Charles C.D. et al.(1991) Biogenic opal in Southern Ocean sediments over the last 450,000 years: Implication to surface water chemistry and circulation, Paleoceanography 6, 697-728.
アルカ リポンプの働き
1.氷期の大気中二酸化炭素濃度減少の謎1.氷期の大気中二酸化炭素濃度減少の謎酸化炭素濃度減少の謎
