| 搶嫗戝妛戝婥奀梞尋媶強 奀梞柍婡壔妛暘栰 |
戝婥CO2偑彮側偐偭偨昘婜偺奀 |

| 搶嫗戝妛戝婥奀梞尋媶強 奀梞柍婡壔妛暘栰 |
戝婥CO2偑彮側偐偭偨昘婜偺奀 |
| 侾丏昘婜偺戝婥拞擇巁壔扽慺擹搙尭彮偺撲 |
丂抧媴娐嫬偺徻嵶側曄摦婰榐偼昘壨偺昘偺拞偵傂偭偦傝偲巆偝傟偰偄偨丅儃乕儕儞僌偱昘彴偵寠傪孈偭偰備偔偲丄怺偔側傞偵偮傟偰傛傝愄偵偱偒偨昘偑庤偵擖傞丅偦偺巁慺傗悈慺偺摨埵懱斾偺暘愅偐傜丄昘偑偱偒偨偲偒偺抧昞柺偺暯嬒婥壏偑悇掕偱偒傞偙偲偼侾俋俇侽擭戙偐傜僐儁儞僴乕僎儞戝妛偺僟儞僗僈乕僪傜偺尋媶偱抦傜傟偰偄偨丅昘偺拞偵偼婥朅傗僟僗僩傕娷傑傟偰偍傝丄偦傟傜偺暘愅偐傜愄偺娐嫬傪撉傒庢傞偙偲偑偱偒傞偐傕抦傟側偄丅傑偢丄嬻婥偑暵偠崬傔傜傟偰偄傞偼偢偱偁傞偐傜丄曄幙傪偆偗側偄傛偆偵偆傑偔庢傝弌偣傟偽戝婥慻惉偺曄摦偑傢偐傞偩傠偆丅偟偐偟丄傢偢偐側婥朅拞偺擇巁壔扽慺傗儊僞儞側偳婥懱惉暘偵偮偄偰偼傑偩偦偺應掕庤朄偵栤戣偑偁偭偨丅侾俋俉侽擭戙偵擖偭偰傛偆傗偔偦偺婰榐傪撉傒弌偡偙偲偵僼儔儞僗傗僗僀僗偺尋媶幰偑惉岟偟丄傑偢嶻嬈妚柦慜偼戝婥拞偺擇巁壔扽慺擹搙偼俀俉侽ppm偱偁偭偨偺偵懳偟丄嵟廔昘婜偵偼俀侽侽ppm慜屻偱栺俉侽ppm傕掅偐偭偨偙偲偑撿嬌僶乕僪婎抧偺昘彴僐傾乕偺暘愅偐傜柧傜偐偵偝傟偨丅堷偒懕偄偰丄撿嬌儃僗僩乕僋婎抧偺昘彴僐傾乕偐傜偼丄夁嫀係俀枩擭偵傢偨傞戝婥拞偺擇巁壔扽慺傗儊僞儞偑婥壏偲楢摦偟丄偄偮傕昘婜偵偦傟傜偺擹搙偑掅偔側傞廃婜揑曄摦傪偟偰偄傞偙偲傕帵偝傟偨[1]丅昘婜偵偼棨忋怉暔偑尭彮偟偰偄偨偙偲偼柧傜偐側偺偱丄偦偺暘偺扽慺偲戝婥偐傜尭彮偟偨擇巁壔扽慺偼偄偢傟傕奀梞偵媧廂偝傟偰偄偨偼偢偱偁傞丅偙偺偙偲偼怺奀掙懲愊暔拞偺桳岴拵偺扽慺摨埵懱斾偑昘婜偵兟侾俁C抣偵偟偰暯嬒侽丏俁俆侎傎偳晧偺懁偵偢傟偰偄傞乮偮傑傝棨忋怉暔偺寉偄扽慺偑奀梞偵壛傢偭偨乯偙偲偵傛偭偰傕巟帩偝傟偰偄傞丅
丂偩偑丄偳偺傛偆偵偟偰昘婜偺奀偑夁忚偺擇巁壔扽慺傪庢傝崬傫偩偺偩傠偆偐丅
| 俀丏梟夝億儞僾 |
丂偦偺尨場偲偟偰傑偢摢偵晜偐傇偙偲偼丄昞憌悈壏偺掅壓偵傛傞擇巁壔扽慺偺梟夝搙偺憹壛偱偁傞丅栚偵偼尒偊側偄偑丄戝婥偲奀梞昞憌偺娫偱偼奀柺傪捠偟偰忢偵婥懱偺岎姺偑峴傢傟偰偄傞丅
丂奀悈偺応崌丄婥懱偺梟夝搙偼暔棟壔妛揑暯峵偵傛偭偰悈壏偑壓偑傞偲傢偢偐側偑傜憹偟丄墫暘偑擹偔側傞偲媡偵尭彮偡傞丅昘婜偵偼奀梞昞憌偺悈壏偑掅偐偭偨偺偱丄戝婥拞偺擇巁壔扽慺偑傛傝懡偔奀偵梟偗崬傒丄擹搙傪尭彮偝偣偨偺偐傕抦傟側偄丅偙偺儊僇僯僘儉傪乽梟夝億儞僾乿偲偟傛偆丅
丂偦偺岠壥偼丄昘婜偺奀梞娐嫬偑偳傫側忬懺偱偁偭偨偐偵傛偭偰寛傑傞丅嵟廔昘婜偺昞柺奀悈偺壏搙偼尰嵼傛傝俀亷慜屻掅偐偭偨偙偲偑怺奀懲愊暔拞偺桳岴拵壔愇側偳偺朙晉側尋媶偐傜抦傜傟偰偄傞丅堦曽丄戝棨昘彴偑惉挿偟偨偙偲偵傛偭偰奀悈弨偑栺侾俀侽m傎偳掅壓偟偨丅偦偺偨傔偵尭彮偟偨奀悈検偐傜丄憹壛偟偨墫暘傕寁嶼偱偒傞丅偦偺寢壥偼丄椉曽偺岠壥偑憡嶦偟偰丄戝婥拞偺擇巁壔扽慺擹搙傪崅乆侾侽ppm掱搙尭彮偝偣傞偵偡偓側偄偙偲偑敾柧偟偨丅梟夝億儞僾偱偼偲偰傕栺俉侽ppm傕偺尭彮傪愢柧偡傞偺偼柍棟側偺偱偁傞丅偟偨偑偭偰丄奀梞拞偺暔幙弞娐偵偍偗傞惗暔揑偁傞偄偼壔妛揑曄壔側偳丄懠偺梫場傪峫偊側偗傟偽側傜側偄丅
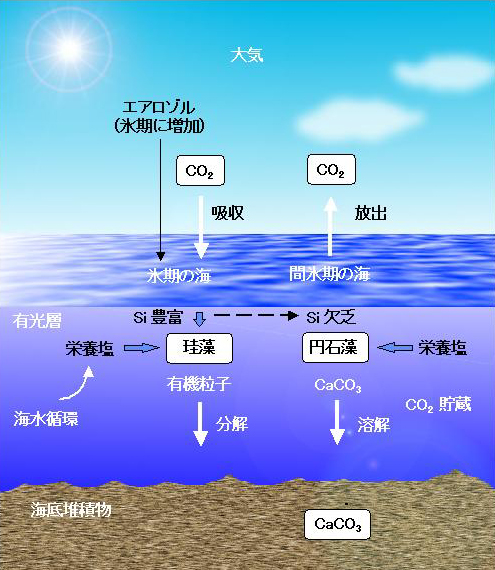
丂恾侾偼丄杒懢暯梞偵偍偗傞梟懚柍婡扽慺擹搙偺墧捈暘晍傪偦傟偧傟偺巟攝梫場偵暘偗偰帵偟偨傕偺偱偁傞[2]丅怺憌悈拞偺擹搙偼昞憌悈偵斾傋偰栺俀侽亾夁忚偵側偭偰偄傞丅偙傟偼奀梞昞憌偱岝傪棙梡偟偰怉暔僾儔儞僋僩儞偑偮偔偭偨桳婡暔傗扽巁僇儖僔僂儉偑丄巰屻昞憌偐傜捑崀偟怺憌悈拞偱僶僋僥儕傾偵傛偭偰暘夝丄傑偨偼壔妛揑偵梟夝偡傞偙偲偵傛傞丅偮傑傝丄旝彫側奀梞惗暔偼丄昞憌悈偺梟懚柍婡扽慺傪懱撪偵庢傝崬傒怺憌傊塣傫偱曻弌丒挋憼偡傞丄偄傢偽億儞僾偺栶妱傪壥偨偟偰偄傞偙偲偵側傞丅昘婜偵偼丄偙傟傜惗暔偺摥偒偵傛偭偰乮偨偲偊偽丄惗暔惗嶻検傗扽巁僇儖僔僂儉偺惗惉丒梟夝夁掱偑曄傢傞側偳偟偰乯丄尰嵼傛傝懡検偺扽慺偑庡偵奀梞怺憌偵曐帩偝傟偰偄偨偲巚傢傟傞丅偦偺偝傑偞傑側壜擻惈傪偮偓偵峫偊偰傒傛偆丅
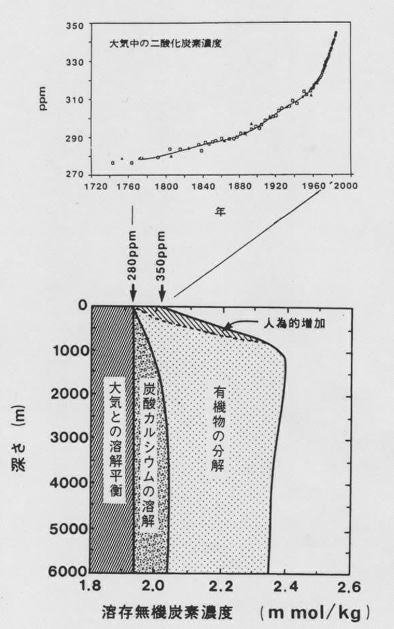
恾侾丏杒懢暯梞偵偍偗傞梟懚柍婡扽慺偺暘晍偲偦偺擹搙傪巟攝偡傞場巕丏
| 俁丏惗暔億儞僾 |
丂奀梞偺惗暔惗嶻偼岝偺撏偔奀梞昞憌乮桳岝憌乯偱峴傢傟傞偑丄僾儔儞僋僩儞偼巰屻丄桳婡暔棻巕偲偟偰怺奀傊塣偽傟偰昞憌偐傜彍偐傟傞偨傔丄偦偺晄懌偟偨梟懚柍婡扽慺偺曗偆宍偱戝婥偐傜擇巁壔扽慺偑梟偗崬傓丅偙偺夁掱傪乽惗暔億儞僾乿偲屇傫偱偄傞丅偦偺斀墳偼
.jpg)
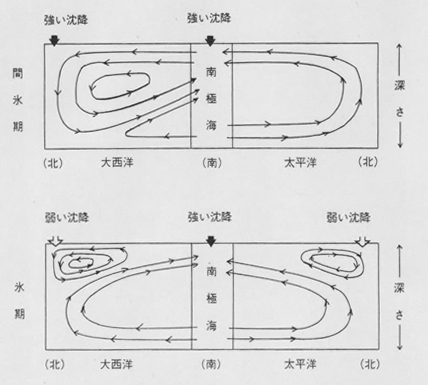
恾俀丏昘婜偲娫昘婜偺奀梞弞娐偺堘偄[2]丏
| 係丏傕偆堦偮偺旝検昁恵尦慺丗揝 |
丂尰嵼丄拞堒搙奀堟偺昞憌悈偼怉暔僾儔儞僋僩儞偵棙梡偝傟偰塰梴墫偑傎傏姰慡偵屚妷偟偰偄傞偑丄儕儞傗拏慺偑巆偭偰偄側偑傜惗暔惗嶻偑偁傑傝崅偔側傜側偄奀堟偑偁傞丅搶戝暯梞偺愒摴晅嬤傗撿嬌廃曈側偳偱偁傞偑丄偦傟傜偺奀堟偱偼丄嵟嬤旝検昁恵尦慺偺堦偮偱偁傞揝偑晄懌偟偰偄傞偨傔偵惗暔惗嶻偑惂尷偝傟偰偄傞偙偲偑柧傜偐偵側偭偰偒偨丅儕乕價僢僸偺嵟彮棩偱抦傜傟傞昁恵塰梴慺偺拞偵丄僇儕僂儉傗儅僌僱僔僂儉側偳偲暲傫偱揝偑偁傞丅揝偼僋儘儘僼傿儖偺峔惉尦慺偱怉暔僾儔儞僋僩儞偺憹怋偵晄壜寚偱偁傞丅偟偐偟丄奀悈拞偱偼揝偼庡偵俁壙偱懚嵼偟悈巁壔暔傪宍惉偡傞偨傔丄旕忢偵晄埨掕偱暯嬒懾棷帪娫偑抁偄丅偦偺偨傔奀悈拞偵朙晉側僇儕僂儉傗儅僌僱僔僂儉偲偼堎側傝丄揝偼應掕偡傞偺偑崲擄側傎偳擹搙偑掅偄丅僇儕僼僅儖僯傾偺儌僗丒儔儞僨傿儞僌尋媶巤愝偺僕儑儞丒儅乕僠儞偑僋儕乕儞媄弍傪嬱巊偟偰暘愅偟偨寢壥丄昞憌奀悈拞偺揝偺擹搙偼侾僫僲儌儖乮侾侽-9儌儖乯埲壓偱偁傝丄偲偔偵儕儞巁傗徤巁偺傛偆偵偲偔偵昞憌偱掅偄偙偲偑傢偐偭偨丅偦傟傜偺奀堟偱奀悈傪嵦庢偟丄彮検偺揝傪壛偊傞偲妶敪偵怉暔僾儔儞僋僩儞偑憹怋偡傞偙偲偐傜丄惗暔惗嶻偺惂尷梫場偲側偭偰偄傞偙偲偑悇掕偝傟偨丅
丂傕偟昘婜偵偦偺傛偆側奀堟偵揝偑嫙媼偝傟偨偲偡傟偽丄惗暔億儞僾偺岠棪傪偨偐傔戝婥拞偺擇巁壔扽慺擹搙傪尭彮偝偣偨偙偲偑婜懸偝傟傞丅恾俁偵帵偡昘彴僐傾乕拞偺僟僗僩偺検偑昘婜偵寖憹偡傞偙偲偱傕傢偐傞傛偆偵丄昘婜偵偼婥壏偑掅偔偔姡憞婥岓偩偭偨偨傔丄搚忞棻巕側偳偑晳偄忋偑傝傗偡偄忬懺偩偭偨丅偦偺僟僗杕偵娷傑傟偨揝偑戝婥傪捠偟偰尰嵼偺晉塰梴掅僋儘儘僼傿儖奀堟偵塣偽傟丄奀悈偵梟弌偟偰惗暔惗嶻傪崅傔偨偙偲偑梕堈偵憐憸偝傟傞偺偱偁傞丅
丂偟偐偟側偑傜丄偙傟傜惗暔億儞僾偺儊僇僯僘儉偺徻嵶偼偲傕偐偔丄惗暔惗嶻偺曄壔偵敽偆桳婡暔偺怺憌傊偺僼儔僢僋僗偺憹壛偱戝婥拞偺俉侽ppm偺尭彮傪愢柧偟傛偆偲偡傞帋傒偵偼偳偆偟偰傕墇偊傜傟側偄戝偒側僴乕僪儖偑偁偭偨丅偦傟偼丄怺憌悈拞偱偺桳婡暔偺巁壔暘夝偵偼梟懚巁慺偑巊傢傟傞偑丄偁傑傝偵桳婡暔棻巕偺捑崀偑懡偔側傞偲怺憌悈偺巁慺偑徚旓偟偮偔偝傟偰巁寚忬懺偵側偭偰偟傑偆偲偄偆偙偲偱偁傞丅偙傟偼丄怺奀偺憡摉晹暘偑僿僪儘忬懺偵側傞偙偲傪堄枴偟丄昘婜偵傕尨惗摦暔偱偁傞掙惐桳岴拵偑惗偒偰偄偨偙偲傪帵偡怺奀懲愊暔偺婰榐偲柧傜偐偵柕弬偡傞丅
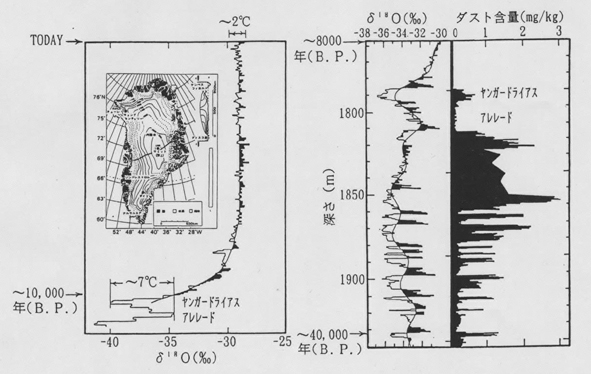
恾俁丏僌儕乕儞儔儞僪偺僉儍儞僾僙儞僠儏儕乕乮嵍乯偲僟僀俁乮塃乯偺昘彴僐傾偺巁慺摨埵懱斾偍傛傃僟僗僩娷検偺曄摦婰榐丏
| 俆丏傾儖僇儕億儞僾偺摥偒 |
丂偦偙偱巆傞壜擻惈偼丄扽巁僇儖僔僂儉偺惗惉偲梟夝偺僶儔儞僗偑曄傢傞偙偲偵傛偭偰戝婥拞偺擇巁壔扽慺偑奀偵媧廂偝傟偨偺偱偼側偄偐偲偡傞峫偊偱偁傞丅僒儞僑偑擇巁壔扽慺偺媧廂尮偲偄偆恖偵偼摢傪椻傗偟偰峫偊偰偄偨偩偔偙偲偲偟偰丄偦偺擇巁壔扽慺媧廂偺尨棟偼拞榓斀墳偱帵偝傟丄梟懚巁慺偼娭學偣偢丄傾儖僇儕搙偑憹壛傪偡傞丅偟偨偑偭偰傾儖僇儕億儞僾偲屇傇偑丄偙偺夁掱偼丄怺奀偑夁忚偺扽慺傪挋憼偟偰傕柍巁慺忬懺偵側傜偢偵偡傓崱偺偲偙傠桞堦偺夝寛嶔側偺偱偁傞丅
丂奀梞昞憌偺奀悈偼扽巁僇儖僔僂儉偵懳偟偰夁朞榓偺忬懺偵偁傝丄桳岴拵丄墌愇憯丄僒儞僑側偳偺惗暔偑扽巁僇儖僔僂儉傪惗惉偡傞丅偮傑傝丄忋婰偺斀墳偑塃偐傜嵍傊恑傓丅堦曽丄怺奀偱偼埑椡偑偐偐傝扽巁僇儖僔僂儉偺梟夝搙偑憹偡偙偲傗桳婡暔偺暘夝偺偨傔偵擇巁壔扽慺偺暘埑偑崅偔側傞偙偲偐傜丄偁傞怺搙傪墇偊傞偲枹朞榓偵側傝丄捑崀偟偰偒偨僾儔儞僋僩儞偺扽巁僇儖僔僂儉焩偼梟夝偡傞丅昞憌奀悈偺傾儖僇儕搙偑昘婜偵崅偐偭偨偙偲偼丄擇巁壔扽慺偺戝婥偲奀悈娫偺暔棟揑側梟夝暯峵偐傜寁嶼偱媮傔傞偙偲偑偱偒傞丅恾係偵帵偡傛偆偵嵟廔昘婜偺昞憌奀悈偼丄嶻嬈妚柦慜偵斾傋偰pH偼侽丏侾俆掱搙丄傑偨傾儖僇儕搙偼侾侾侽儅僀僋儘摉検掱崅偐偭偨偙偲偑傢偐傞丅偦偙偱昘婜偵偼側傫傜偐偺棟桼偱丄扽巁僇儖僔僂儉偑傛偔夝偗傞傛偆偵側偭偨偺偱偼側偄偐偲偡傞愢偑弌偝傟偨丅偨偲偊偽儅僒僠儏僙僢僣岺壢戝妛偺E.
A.儃僀儖偵傛傟偽丄惗暔惗嶻偑崅偔側偭偰奀掙偵摓払偡傞桳婡棻巕偺僼儔僢僋僗偑憹戝偟丄偦偺暘夝偵傛偭偰
惗偠偨擇巁壔扽慺偑奀掙偺 扽巁僇儖僔僂儉 傪梟夝傪壛懍偡傞偙偲偑峫偊傜傟傞丅偦偺寢壥丄怺憌悈偺傾儖僇儕搙偑憹壛偟丄偦偺奀悈偑奀梞弞娐偵傛偭偰昞憌偵弌偰戝婥偵愙偡傞偲
擇巁壔扽慺傪媧廂偡傞偙偲偵側傞丅嬶懱揑偵偦偺岠壥傪榑偠偨榑暥傕偦偺屻偄偔偮偐敪昞偝傟偰偄傞丅偟偐偟丄偨偲偊偙偺傛偆偵怺奀掙偱
扽巁僇儖僔僂儉 偺梟夝偑憹偊偨偲偟偰傕丄偦偺塭嬁偑戝婥偵尰傟傞偵偼奀梞弞娐偺帪娫僗働乕儖偐傜峫偊偰彮側偔偲傕悢昐擭偼偐偐傞偵堘偄側偄丅偟偐偟丄昘彴僐傾偺
擇巁壔扽慺擹搙傗揇扽僐傾偺扽慺摨埵懱偑帵偡戝婥 拞偺擇巁壔扽慺擹搙 偺曄摦偼
傢偢偐俀侽乣俁侽 擭偱婲偭偰偄傞偺偱偁傞丅偮傑傝丄偙偺怺奀掙扽巁墫梟夝愢偩偗偱愢柧偡傞偺偵偼柍棟偑偁傞偲偄偊傛偆丅
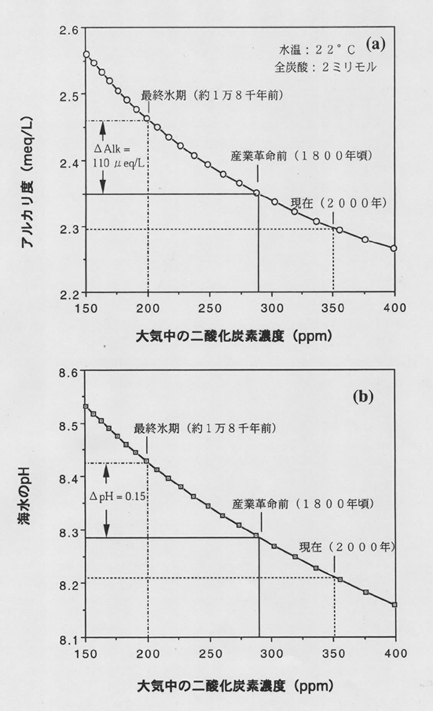
恾係丏戝婥偲暯峵偵偁傞昞憌奀悈偺傾儖僇儕搙乮a乯偲pH乮b乯丏
| 俇丏僔儕僐儞壖愢 |
丂偦偆偙偆偡傞拞偱栰嶈偼怴偟偄夝寛嶔傪巚偄偮偒丄戝応偲嫟摨偱榑暥偵敪昞偟偨[3]丅偮傑傝丄怺奀偱偺扽巁僇儖僔僂儉偺梟夝偑憹偊偨偺偱偼側偔丄偦偺焩傪傕偮僾儔儞僋僩儞偺憹怋偑梷惂偝傟偨偺偱偼側偄偐偲峫偊偨偺偱偁傞丅昞憌奀悈偵儕儞傗拏慺側偳懠偺塰梴墫偑偁傟偽丄梟懚働僀慺偑偁傞応崌偵偼僆僷乕儖乮柍掕宍働僀巁墫乯偺焩傪傕偮宂憯偑桪愭揑偵斏怋偡傞偙偲偼埲慜偐傜抦傜傟偰偄偨丅働僀慺偑寚朢偟偰側偍懠偺塰梴墫偑巆偭偰偄傞応崌偵墌愇憯偑憹怋偡傞丅塰梴墫偺扗偄崌偄偱偼丄扽巁僇儖僔僂儉偺妅傪帩偮墌愇憯偼側偤偐働僀憯偵堷偗傪庢傞偲偄偆帺慠奅偺愛棟偑偁傞丅偦偺傛偆側忦審壓偱偼丄墌愇憯偼昁梫側拏慺傗儕儞傪宂憯偵扗傢傟偰憹怋偟偵偔偄偺偱偼側偄偐偲峫偊偨偺偱偁傞丅昘婜偵偼奀梞昞憌傊塣偽傟傞僄傾儘僝儖偺検偑憹戝偟丄偦偙偐傜梟偗弌偟偨梟懚働僀慺偼宂憯偺憹怋傪懀恑偟偨偼偢偱偁傞丅丂丂丂丂丂丂丂
丂偟偨偑偭偰丄宂憯偑戩墇偡傞婜娫偑挿偔側傞偲偐丄偁傞偄偼宂憯偑塰梴墫傪巊偄愗偭偰偟傑偭偰墌愇憯偑惉挿偱偒側偄側偳偺尰徾偑婲偙傟偽丄(2)
幃偵徠傜偟偰傾儖僇儕搙偼忋徃偟丄戝婥 偐傜擇巁壔扽慺 傪媧廂偱偒傞偙偲偵側傞丅怴偨側曗懌傪壛偊偰偦偺壖愢傪弎傋傞偲師偺傛偆偵側傞乮恾俆嶲徠乯丅
恾俆丏僔儕僐儞桿摫扽慺弞娐儌僨儖偺奣棯丏
丂乮侾乯僇僫僟杒曽偺昘壨曵夡乮僴僀儞儕僢僸丒僀儀儞僩偵懳墳乯偱杒戝惣梞昞憌偵扺悈偑棳擖偟丄擬墫弞娐乮僽儘僢僇乕偺僐儞儀儎乕儀儖僩乯傪掆巭偟偦偆側傎偳庛傔偨丅偦偺寢壥丄杒戝惣梞偱偺昞憌悈偑掆懾偟偰捑崀偟側偔側傝丄儊僉僔僐榩棳偼杒忋偱偒側偔側偭偰儓乕儘僢僷偑媫懍偵姦椻壔偡傞丅
丂乮俀乯姦椻壔偵傛偭偰丄姡憞婥岓偲側偭偨戝抧偐傜搚忞棻巕偑晳偄忋偑傝傗偡偔側傝丄奀梞昞憌傊偺僄傾儘僝儖偺崀壓検偑憹偟偨丅偮傑傝丄僄傾儘僝儖偵傛偭偰傕偨傜偝傟偨
働僀慺 偑奀梞昞憌偵朙晉偵側傝丄宂憯偺憹怋傪懀恑偡傞偲摨帪偵丄扽巁僇儖僔僂儉偺妅傪傕偮墌愇憯偺憹怋傪梷惂偟偨丅
丂乮俁乯偦偺寢壥丄昞憌奀悈偵偍偗傞 扽巁僇儖僔僂儉 偺惗惉検偑憡懳揑偵尭彮偟丄捑崀棻巕偺桳婡扽慺/
扽巁僇儖僔僂儉 斾傪墴偟忋偘傞丅偦偟偰奀悈偺傾儖僇儕搙傪憹壛偝偣傞偲偲傕偵pH
偑崅偔側偭偰戝婥偐傜擇巁壔扽慺傪媧廂偟偨丅
偙偺傛偆偵奀梞昞憌偱偺夁掱側傜偽丄俀侽乣俁侽擭偱戝婥
拞偺擇巁壔扽慺擹搙 傪曄壔偝偣傞偙偲傕壜擻偩傠偆丅惗暔偺庬偺岎戙偵偼偙偺傛偆偵堦掕偺儖乕儖偑偁傞応崌偑偁傞丅偙偺惗暔儕儞働乕僕傪捠偟偰丄壔妛揑偵偼撈棫偟偨斀墳偱偁傞扽巁僇儖僔僂儉
偺惗惉丒梟夝僒僀僋儖偲 僔儕僇偺僒僀僋儖偑帺慠奅偱偼岻柇偵寢傃偮偄偰偄傞傛偆偱偁傞丅
| 俈丏怴偨側徹嫆扵偟 |
丂嵟嬤偺儌僨儖寁嶼偱偼丄慡奀梞偱惗嶻偝傟傞扽巁僇儖僔僂儉偑係妱尭彮偡傟偽丄僔儕僐儞壖愢偺儊僇僯僘儉偱昘婜戝婥偺擇巁壔扽慺擹搙偺愢柧偑壜擻偲偄傢傟傞丅墌愇憯偲宂憯偺庬偺岎戙偼丄儕儞丄拏慺丄揝側偳偵懳偟偰梟懚働僀慺偺嫙媼偑憡懳揑偵晄懌偟偨奀堟偱幚嵺偵婲偙傝偆傞丅杒戝惣梞丄愒摴戝暯梞傗撿嬌奀偺撿堒係俆乣俆侽搙埲杒偱偼丄梟懚働僀慺偲徤巁偺斾偑宂憯偑昁梫偲偡傞侾埲壓偱偦偺岓曗奀堟偲偄偆偙偲偵側傞丅嵟嬤丄僐儘儞價傾戝妛儔儌儞僩抧媴娤應尋媶強偺C.D.僠儍乕儖僘傜偑撿嬌廃曈奀堟偺怺奀懲愊暔偺巁慺摨埵懱斾偲偲傕偵僆僷乕儖偲扽巁僇儖僔僂儉娷検傪徻偟偔敪昞偟偰偄傞偑丄偦偺堦椺傪恾俇偵帵偟偨丅懲愊暔拞偺僆僷乕儖娷検偼丄奀悈傪捑崀拞偁傞偄偼奀掙偱杽杤偡傞傑偱偺娫偵梟夝偝傟偢偵巆偭偨傎傫偺堦晹暘偵偡偓側偄偺偱丄偦偺梟夝偲曐懚偵娭偡傞條乆側夁掱偑曄傢傟偽塭嬁偝傟傞丅偟偐偟丄僠儍乕儖僘傜[4]偼丄條乆側専摙傪峴偭偨屻丄僆僷乕儖娷検偼庡偵奀梞昞憌偱偺惗暔惗嶻傪昞偟偰偄傞傕偺偲寢榑偟偰偄傞丅摨條偺壖掕偼丄扽巁僇儖僔僂儉偵偮偄偰傕惉傝棫偮偱偁傠偆丅
丂恾俇偐傜柧傜偐側傛偆偵丄夁嫀栺侾枩擭偺娫偼扽巁僇儖僔僂儉偑戩墇偟偰偄傞偑丄侾枩俋愮擭偐傜俀枩俆愮擭偺嵟廔昘婜偺帪戙偵偼扽巁僇儖僔僂儉偼悢亾偵傑偱屻戅偟丄宂憯偑庡偵側傞偙偲偑傢偐傞丅宂憯偲墌愇憯偺庬偺岎戙偑婲偭偰偄傞偙偲偼丄恾俈偵帵偡僆僷乕儖偲扽巁墫偺偒傟偄側媡憡娭娭學偐傜傕悇掕偱偒傞丅傑偨丄夁嫀侾枩擭偺娫偼栺俋侽亾偑惗暔惈扽巁墫偲僆僷乕儖偱愯傔傜傟偰偄傞偑丄嵟廔昘婜偵偼俀侽乣俀俆亾偱偦偺懠偼棨偐傜塣偽傟偨擲搚峼暔側偳偱偁傞丅懲愊暔偺擭戙偐傜棨婲尮旝彫棻巕偺懲愊懍搙傪寁嶼偡傞偲丄昘婜偺曽偑尰嵼偺娫昘婜傛傝侾寘戝偒偄丅昘婜偵業弌偟偨棨扞偐傜偼偙偽傟偨傕偺傕娷傑傟傞偐傕偟傟側偄偑丄戝晹暘偼戝婥傪宱桼偟偰塣偽傟偨傕偺偲巚傢傟傞丅
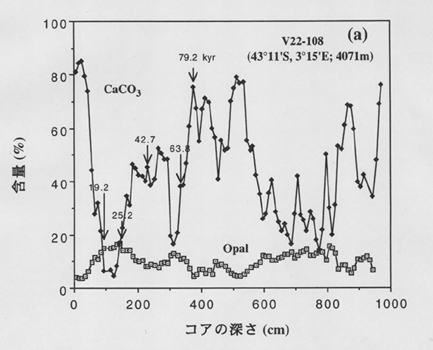
恾俇丏撿戝梞怺奀僐傾偺扽巁僇儖僔僂儉偲僆僷乕儖娷検偺曄摦[5]
恾拞偺悢抣偼愮擭偺扨埵偺擭戙傪昞偡丏
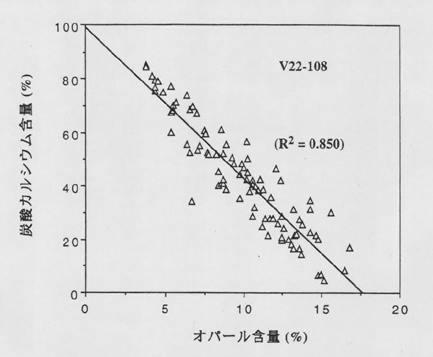
恾俈丏V俀俀亅侾侽俉僐傾偺扽巁僇儖僔僂儉偲僆僷乕儖娷検偺娭學丏
嶲峫暥專丗
[1] Petit J. R. et al. (1999), Climate and atmospheric
history of the past 420,000 years from the Vostok ice core, Antarctica.
Nature 399, 429-436.
[2] 栰嶈媊峴 (1994)丄抧媴壏抔壔偲奀丄搶嫗戝妛弌斉夛丄p196.
[3] Nozaki, Y. and T. Oba (1995) Dissolution of
calcareous tests in the ocean and atmospheric carbon dioxide, In: Biogeochemical
Processes and Ocean Flux in the Western Pacific. H. Sakai and Y. Nozaki,
eds., TerraPub., Tokyo, p83-92.
[4] Charles C.D. et al.(1991) Biogenic opal in Southern
Ocean sediments wver the last 450,000 years: Implication to surface water
chemistry and circulation, Paleoceanography 6, 697-728.
![]()